


生物科学専攻(植物学系)・准教授 竹中 瑞樹

RNA編集とはRNAの特定の位置で塩基の置換や、挿入、欠失がおこる現象である。陸上植物オルガネラのRNA編集は、原生動物のミトコンドリアや、哺乳類の核に続き、約30年前に発見された。植物のミトコンドリアや葉緑体では特定のC(シチジン)が選択されU(ウリジン)に変換される。その結果、翻訳産物であるタンパク質のアミノ酸配列も変化する。つまりこの機構は、遺伝子産物を質的に制御する機構といえる。
われわれは植物オルガネラのRNA編集機構に関与するタンパク質因子を、次々と同定してきた。これによって、植物オルガネラが他の生物とは全く異なるRNA編集機構を持つことが明らかになってきた。数百を超えるRNA編集部位は、約35アミノ酸からなる保存された配列を10−20個繰り返しもつ、PPRタンパク質(pentatricopeptide repeat protein)により特異的に認識される。最近の研究では、PPRの各モチーフがRNAを一塩基ずつ認識することにより、各RNA編集位を特異的に認識することが明らかになってきた(図1)。またPPRタンパク質と複合体を形成して、数十を超えるRNA編集部位を同時に制御するMORF(Multiple Organellar RNA editing factor)も単離した。RNA編集に関与するこれらの多様な因子の発見は、植物オルガネラにおいて複雑なRNA編集複合体が形成されることを示唆している(図1)。現在われわれはRNA編集反応の分子機構および、MORFを含むRNA編集複合体の全体像解明を目指して解析をすすめている。
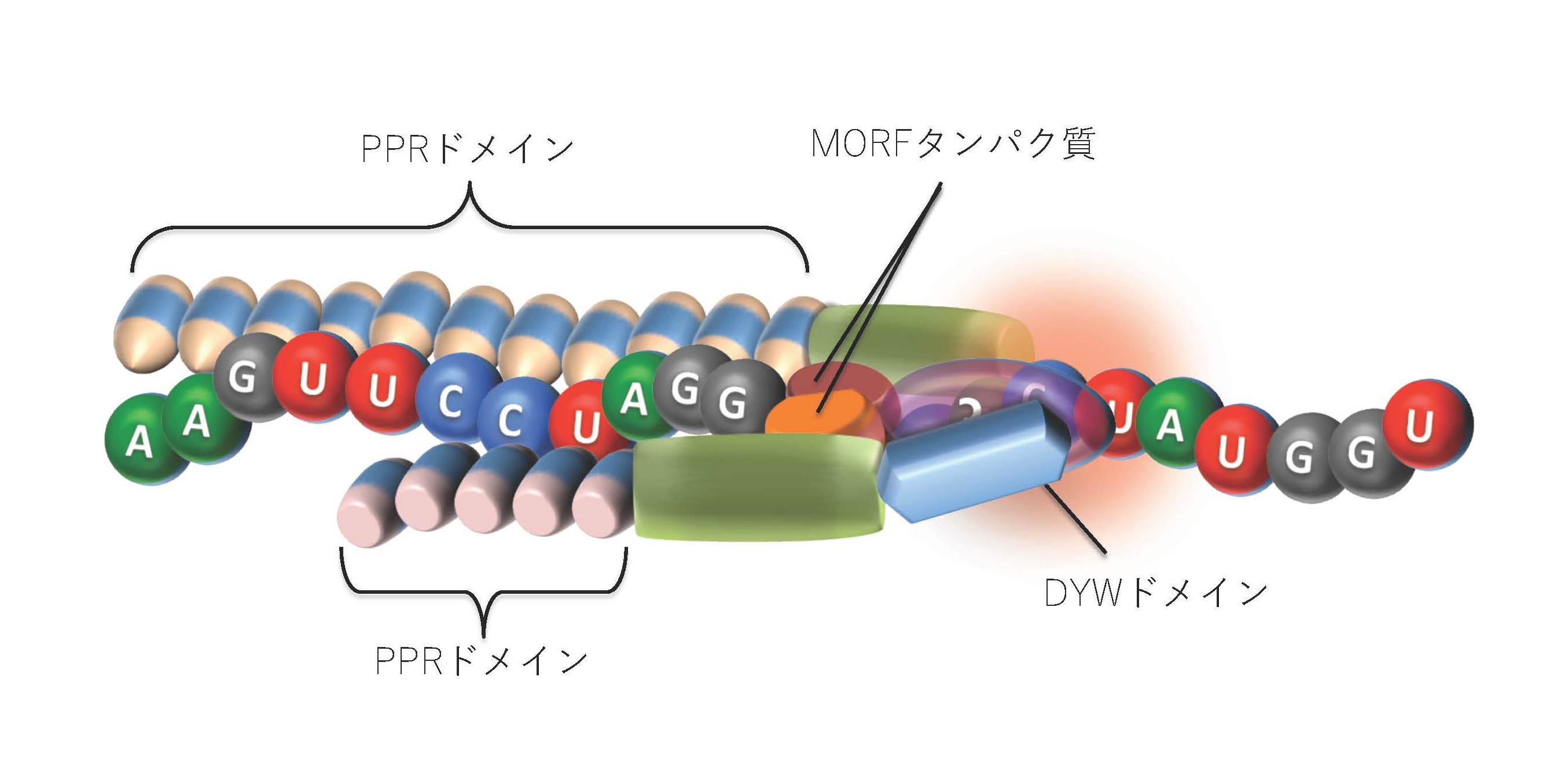
PPRタンパク質の各モチーフはRNA編集部位の近傍の配列を認識する(図上部)。このPPRタンパク質はC末端にRNA編集酵素と考えられるDYWドメインを直接保持する場合もある。しかし、これを保持しない場合にはDYWドメインをもつがRNA結合能は低い他のPPRタンパク質(図下部)と複合体を形成する。このRNA編集複合体形成にはMORFタンパク質のホモまたはヘテロ二量体やその他のタンパク質が関与している。